Genetic basis of morphology and adaptation in Felidae and other mammalian species

Understanding the genetic basis of phenotypic variation is of tremendous importance for uncovering the mechanisms of species’ diversification and evolution. In particular, coat color and pattern are prominent morphological features in mammals and play an essential role in adaptation and survival. Felidae, or the cat family, consists of 36 to 40 species depending on taxonomic classification systems, and exhibits the most diverse fur color, pattern and density of all terrestrial carnivores. Felids also vary greatly in weight, body size, and tail length and morphology. We take the advantage of the latest advances in feline genomics and high-throughput sequencing to unravel the genetics underlying the inherited traits of domestic cats and their wild cousins.
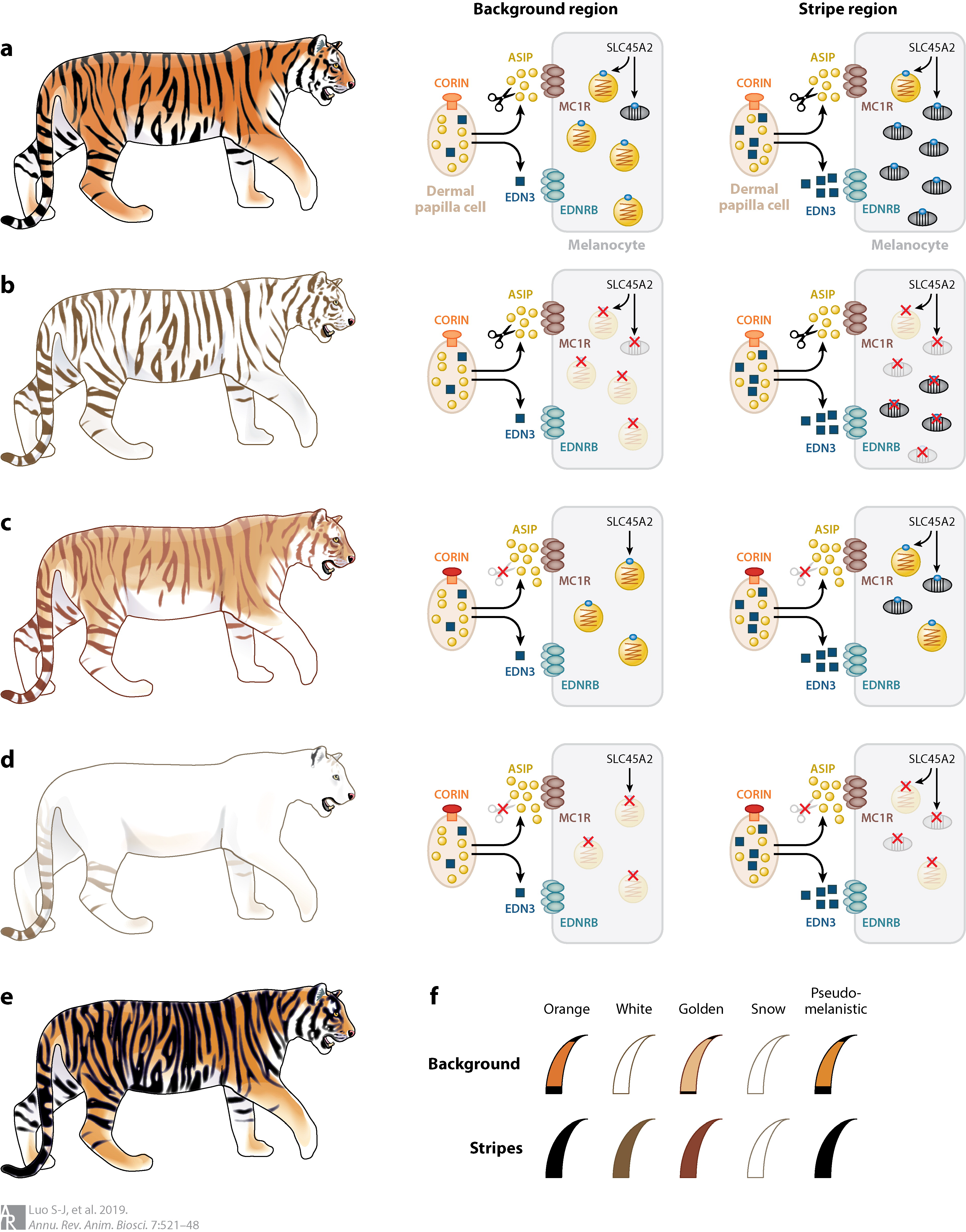
The tiger is generally recognized for its dark stripes against an orange fur, with a few variants found in Bengal tigers, including the charismatic white tiger and the less known golden tabby and nearly stripeless snow tigers. Captive breeding records have suggested that the white and golden tiger coat colors are determined by two independent autosomal recessive loci, however, the precise genetic mechanisms remain elusive until now. We employed a pedigree-based genome-wide association study (GWAS) approach in combination of whole genome resequencing to reveal the causative mutation of white tiger to be a single amino acid change (A477V) in a transporter protein SLC45A2 (Xu et al. 2013), and that of the golden tiger to be H587Y in the transmembrane serine protease CORIN (Xu et al. 2017). A dual homozygote carrying both mutations results in the nearly complete loss of melanin in the snow tiger. We further conducted immunochemistry study to show that the H587Y mutation impairs the function of CORIN in degrading agouti signaling protein, strengthening the inhibitory effect of agouti signaling protein on eumelanogenesis during hair growth cycles and resulting in a prolonged pheomelanin band in the golden tiger’s hair. We also argue that the SLC45A2 p.A477V substitution itself is not a genetic deformity and that the white tiger morph is a viable, perhaps neutral, genetic polymorphism. Despite its low frequency, this polymorphism has survived in the wild for at least several centuries and should be considered a part of the genetic diversity of tigers that is worth conserving (Luo and Xu, 2014). Decoding the genetic basis of tiger pelage color variation has advanced our understanding of mammalian coat color genetics and supported the use of nonmodel species in elucidating the genetic basis of animal adaptive diversity (Xu and Luo, 2014; Luo et al., 2019).
The cat was domesticated around 10,000 years ago. With the variable artificial selection of human preferences, various traits were selectively bred, resulting tremendous variations in modern domestic cats. Thus, the domestic cat serves as an ideal model for an enhanced understanding of the genomic diversity and evolution in Felidae. Using domestic cat model, we have elucidated the genetic basis of the tail-length polymorphism (Xu et al., 2016). Multiple projects are going on with regard to various morphological polymorphisms in domestic and wild cats, such as the silver coat color, dominant white and white spotting coat color?kinked/short tails, and catnip response etc.. In addition, we also explored the genetic basis of coat color variation in domestic yaks with a candidate gene approach (Zhang et al., 2014).